Following some earlier blogs about recently acquired collections I present to you here the Treat collection. This collection was assembled by Asher E. Treat a researcher at City University of New York and the American Museum of Natural History, also New York. This collection is one of the best in the world for mites associated with Lepidoptera (butterflies and moths). Mites have been found associated with most terrestrial and many aquatic organisms, but when it comes to insect hosts, mites on Coleoptera (beetles) and Hymenoptera (bees and wasps) are clearly the most numerous, diverse, and well-known. Still, Lepidoptera have a variety of associated mites.
The Acarology Collection acquired this collection 4 years ago, some years after Treat’s death. The collection consisted of about 37 slide boxes of exceptionally well labelled microscope slides and half a dozen insect drawers of pinned moths (all labelled as hosts of specific mite specimens). The Lepidoptera are being processed at the Triplehorn Insect collection, while we, the Acarology collection, have been working on processing (mostly databasing) the slides. This is proving to be a major job.

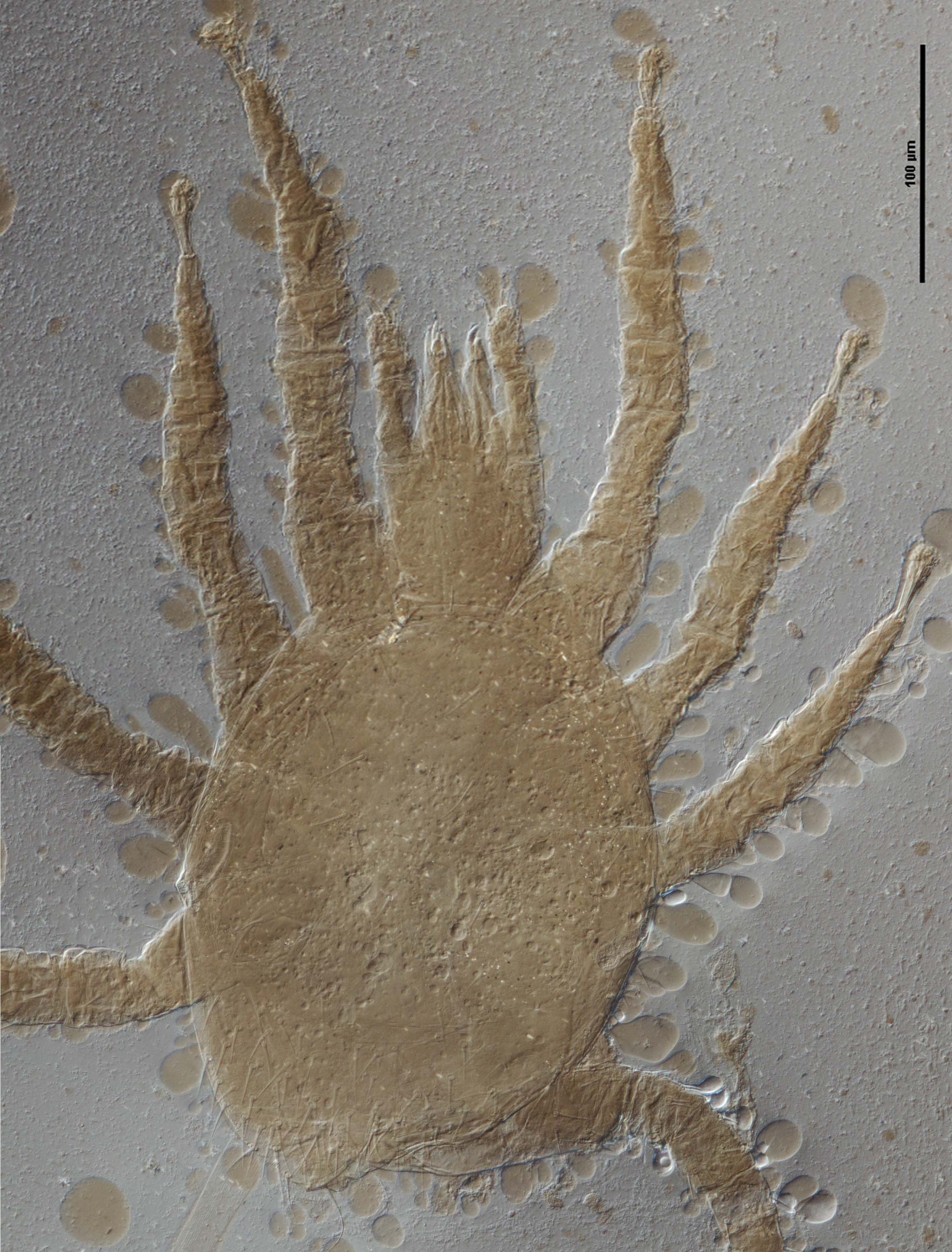
Image of a female of Dicrocheles phalaenodectes, the moth ear mite
Treat got interested in mites associated with moths after finding mites in the ears of noctuid moths. In the process, he figured out the quite amazing life histories of some mites associated with these moths. The most famous is Dicrocheles phalaenodectes, the moth ear mite (family Laelapidae).
These mites break through the tympanic membrane of the ear of the moth and form small colonies inside the ear. By itself not too surprising, but the interesting part Treat discovered was that these mites are always found in one ear only, rarely if ever in both ears. In a way this makes sense. By breaking the tympanic membrane the mites make the moth deaf in that ear. Moths need their hearing to avoid predators (for example bats) so a deaf moth would be easy prey. However, a moth with one functional ear is still able to avoid bats, perhaps not as well as if it had two functional ears, but close enough. Which leaves the question: how do the mites manage to limit infestations to one ear?
Treat did many careful observations and follow-up experiments on this aspect and found that the mites have a very specific set of behaviors ensuring only one ear will be parasitized. The first female to get on a moth (nearly always a fertilized female, the immatures and males do not colonize) crawls to the dorsal part of the thorax, explores a little, after which she proceeds to one ear. Any future colonizers will first go to that same dorsal part of the thorax of the moth and follow the initial female to the same ear. It appears the mites lay a pheromone trail that guides newcomers to the already infested ear, and away from the uninfested one.

Drawing of relative positions of mites in a moth ear
To complete the cycle, young females leaving the ear initially wander around the hosts body (mostly the thorax), congregating around the head at night. They leave the moth by running down its proboscis when it is feeding on flowers. On the flowers, the mites wait for their next host.
Another mite family that is specialized on Lepidoptera, the Otopheidomenidae, is also parasitic, and they will also show up near the ears, but they do not pierce the tympanic membrane, so they do not cause deafness. Unfortunately, we know much less about them, Treat was never able to study their behavior. A range of other mite families have representatives that are regularly found on Lepidoptera, but they are not specialists at the family level: Ameroseiidae, Melicharidae, Erythraeidae, Iolinidae, Cheyletidae, Acaridae, Carpoglyphidae, and Histiostomatidae. That list excludes the occasional “vagrants” that can be found on moths, but that are unlikely to be living on them for extended periods of time. All in all, quite a diverse community.
For those interested in knowing more, Treat wrote a book “Mites of Moths and Butterflies” (1975, Cornell University Press) that is a rare combination of good scholarship (especially natural history) and readability.

Title page of Treat’s book on moth mites
Treat was very careful and noted things like host specimen numbers (if available), which allows current researchers to track down the exact moth from which a given mite came.
This is currently a common approach, but Treat started this in the 1950-ies. And there is more. Based on Treat’s label data we know not only the name of the hosts and the specific locality, but also gender of the host, whether the left or right ear was infested, and the exact part of the body the mites were found on. So we have excellent information, directly from the slides, showing that Proctolaelaps species (family Melicharidae) are nearly always found near the base of the palps [as an aside, Proctolaelaps is a bit of an unfortunate generic name, combining “procto-” = anus and “laelaps” = hurricane; presumably the name

Slide from the Treat collection
refers to a relatively large anal shield]. Such complete data are fantastic for future research, but they also mean a lot more work processing these slides, as every slide has lots of unique data. I want to thank George Keeney, part-time curator of the acarology collection and a series of volunteers, Ben Carey, Rachel Hitt, Mitchell Maynard, Ben Mooney, Jake Waltermyer, and Elijah Williams, for their hard work in accessioning this material.
About the Author: Dr. Hans Klompen is professor in the department of Evolution, Ecology and Organismal Biology and director of the Ohio State University Acarology Collection.