One of the fastest growing areas of concern among scientists is the ever increasing quantity of plastic litter finding its way into aquatic environments. In 2014 alone, approximately 311 million metric tons of plastic were produced, with that number steadily increasing each year (Jonanovic, 2017). Of the plastic produced on a global scale, up to 10% of it ends up in aquatic environments. Specifically, by 2025 the mass ratio of plastic to fish in the oceans is predicted to be 1 to 3 and by 2050 it is expected that plastic will likely surpass fish stocks in the ocean by weight (Jonanovic, 2017). Microplastics, or plastic particles smaller than 5 mm derived from the break down of larger debris or entering the environment directly as microscopic fragments, has been estimated at over 5 trillion floating particles at a weight of 250,000 tons (Sussarellu et al., 2016). While much of the attention of scientists and the public eye have focused on the entanglement in and ingestion of macroplastics by vertebrates, the biological impacts of microplastics on marine organisms is only just now emerging (Wright et al., 2013). The aim of this post is to provide a summary of some of the current research completed looking at the deleterious impacts of microplastics on marine life.

Figure 1: Microplastics collected from the Chesapeake Bay Watershed . https://www.flickr.com/photos/chesbayprogram/16999300502
Given the abundant nature and small dimensions of microplastics, ingestion by marine filter-feeders is of growing cause for concern. Filter-feeding species ingest large volumes of water, and subsequently, large quantities of particles potentially leaving them more susceptible to microplastic pollution. Sussarellu et al. (2016) experimentally exposed reproductively active Pacific oysters (Crassotstrea gigas) to microplastics for two months to access physiological impacts on both adults and offspring. The results of this study demonstrated that adult oysters altered their energy allocation from reproduction to structural growth and maintenance when exposed to microplastics. These alterations lead to reproductive impairment and decreased survival and growth of offspring (figure 2).
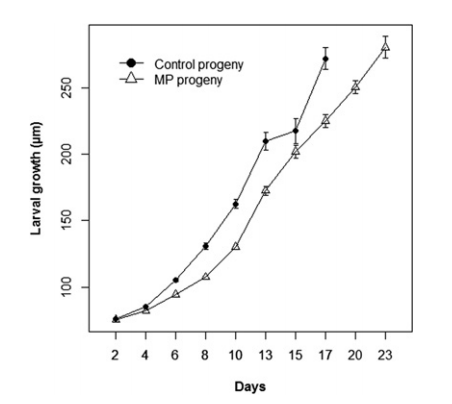
Figure 2: Comparison of larval growth of Pacific oysters between control group and those exposed to microplastics. Suppressed growth of larval oysters exposed to microplastics evident (Sussarellu et al., 2016).
Along with filter feeders, indiscriminate deposit-feeders (aquatic organisms that feed on organic matter that has settled on the sea floor), could be negatively impacted by microplastic pollution. Wright et al. (2013) found that lugworms (Arenicola marina) exposed to natural sediments infused with 5% microplastics depleted energy reserves by up to 50%. The time taken to egest (discharge undigested material) ingested material was 1.5 times longer in lugworms exposed to microplastics compared to control worms. Unfortunately, these two studies represent some of the only research looking at the ecological impacts of microplastic ingestion by filter- and deposit-feeders.
While often considered wholefully insufficiently investigated, recent discoveries regarding the potential negative impacts of microplastic ingestion by fish are emerging throughout the scientific realm (Jovanovic, 2017). Fish could ingest microplastics both intentionally and unintentionally during all stages of development. Laboratory studies focused on on fish ingestion of microplastics have demonstrated varying physiological impacts. Reduced food intake resulting from microplastic gut blockage could lead to decreased energy availability in fish (Mazurais et al., 2015). Rochman et al. (2013) found that marine fish ingesting microplastics sorbed with environmental contaminants bioaccumulate the pollutants and suffer liver toxicity. In the study, fish exposed to microplastics and sorbed contaminants expressed high levels of stress and hepatic (liver) inflammation. These studies indicate the potential for negative physiological impacts on fish populations, but further research is needed to validate these results in the field (Steer et al., 2017).
With potentially devastating physiological implications of microplastic ingestion evident, concentrated efforts on disentangling the ecological impacts of microplastic ingestion on our aquatic ecosystems remains vital. While the elimination of plastic use by humans may be unachievable and the removal of existing microplastics all but impossible, we have a moral obligation to reduce our plastic usage to help alleviate future negative impacts on all aquatic organisms.

Figure 3: https://www.photolib.noaa.gov/htmls/reef2129.htm
References:
Jovanovic B (2017) Ingestion of microplastics by fish and its potential consequences from a physical perspective. Integrated Environmental Assessment and management 13(3): 510-515.
Mazurais D, Ernande B, Quazuguel P, Severe A, Huelvan C, Madec L, Mouchel O, Soudant P, Robbens J, Huvet A, Zambonino-Infante J (2015) Evaluation of the impact of polyethylene microbeads ingestion in Eurpoean sea bass (Dicentrarchus labrax) larvae. Marine Environmental Research 112: 78-85.
Rochman CM, Hoh E, Kurobe T, Teh SJ (2013) Ingested plastic transfers hazardous chemicals to fish and induces heptic stress. Scientific Reports 3(3263): doi.10.1038/srep03263.
Steer M, Cole M, Thompson RC, Lindeque PK (2017) Microplastic ingestion in fish larvae in the western European Channel. Environmental Pollution 226: 250-259.
Sussarellu R, Suquet M, Thomas Y, Lambert C, Fabioux C, Pernet MEJ, Goic NL, Quillien V, Mingant C, Epelboin, Corporeau C, Guyomarch J, Robbens J, Paul-Pont I, Soudant P, Huvet A (2016). Oyster reproduction is affected by exposure to polystyrene microplastics. PNAS. U.S.A. 113, 2430–2435.
Wright SL, Rowe D, Thompson RC, Galloway TS (2013) Microplastic ingestion decreases energy reserves in marine worms. Current Biology 23(23): 1031–1033.
