Normal Physiology of the Human Breast
Prior to sexual maturity, male and female breasts are structurally and functionally similar; they are both comprised of small immature nipples, fatty and fibrous tissue and several duct-like arrangements beneath the areola.
When puberty is underway in males and females, this is where the major structural development occurs. Male breasts remain unchanged due to the lack of high levels of estrogen and progesterone. Females, on the other hand, have significant changes occur due to an assortment of hormones (estrogen, growth hormone, insulin-like growth factor-1, progesterone, and prolactin) (p.739) that cause the female breast to develop into a lactating system.
The mature female breast’s foundational unit is the lobe (each breast contains 15-20) a system of ducts which is comprised of and supported by Cooper ligaments. Each lobe is made up of 20-40 lobules (glands that produce milk). The lobules contain alveolar cells, which are complicated spaces lined with epithelial cells that secrete milk and sub-epithelial cells that contract, passing milk into the arrangement of ducts that leads to the nipple (p. 739).
The lobes and lobules are enclosed and separated by muscle strands and adipose connective tissue which varies in amount depending on weight, genetics, endocrine factors and contributes to the diversity of breast size and shape.
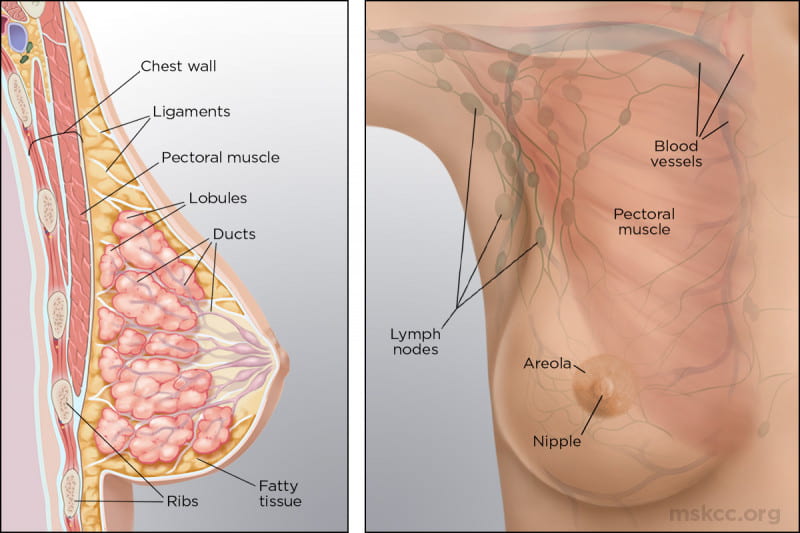
(Memorial Sloan Kettering Cancer Center)
During the reproductive years, breast tissue undergoes cyclic changes in response to hormonal changes of the menstrual cycle. After menopause, adipose deposits and connective tissue increases, glandular breast tissue becomes involuted, and breasts are reduced in size and form. Due to elevated aromatase (decreases circulating estrogen) there can be an increase in white adipose tissue inflammation (p.740).
The function of the female breast is primarily to provide a source of nourishment for the newborn; however, breasts are also a source of pleasurable sexual sensation and in Western cultures have become a sexual symbol (p.741).
(McCance, K. L., & Heuther, S. E. (2019). Pathophysiology: The biological basis for disease in adults and children (8th ed., pp.739-741). St. Louis, MO: Mosby.)
Pathophysiology of Breast Cancer
Except for skin cancer, breast cancer is the most common cancer in American women. Most breast cancer occurs in women older than 50 years. The major risk factors for breast cancer are classified as reproductive, such as nulliparity and pregnancy-associated breast cancer; familial, such as inherited gene syndromes; and environmental and lifestyle, such as hormonal factors and radiation exposure. Some examples of known carcinogenic agents with sufficient evidence in humans that contribute to the development of breast cancer are alcoholic beverages, diethylstilbestrol, estrogen-progestogen contraceptives, estrogen-progestogen menopausal therapy, X-radiation and γ-radiation (Rote, 2019). Other important factors are delayed involution of the mammary gland and increased breast density.
Overall, lifetime risk of breast cancer is reduced in parous women compared to nulliparous women, but pregnancy must occur at a young age. The influence of pregnancy on the risk of cancer also depends on family history, lactation postpartum, and overall parity. Breast gland involution after pregnancy and lactation uses some of the same tissue remodeling pathways activated during wound healing. The presence of macrophages in the involuting mammary gland contributes to carcinogenesis.
Most breast cancers are adenocarcinomas and first arise from the ductal/lobular epithelium as carcinoma in situ. Carcinoma in situ is an early-stage, noninvasive, proliferation of epithelial cells that is confined to the ducts and lobules, by the basement membrane. About 84% of all in situ disease is ductal carcinoma in situ (DCIS); the remainder is mostly lobular carcinoma in situ (LCIS) disease. Ductal carcinoma in situ (DCIS) refers to a heterogenous group of proliferative lesions limited to ducts and lobules without invasion to the basement membrane. DCIS occurs predominantly in women but can also occur in men. DCIS has a wide spectrum of risk for invasive cancers. Preinvasive lesions do not invariably progress to invasive malignancy. Lobular carcinoma in situ (LCIS) originates from the terminal duct-lobular unit. Unlike DCIS, LCIS has a uniform appearance; thus, the lobular structure is preserved. The cells grow in non-cohesive clusters, typically because of a loss of the tumor-suppressive adhesion protein E-cadherin. Also, unlike DCIS, LCIS is found as an incidental lesion from a biopsy and not mammography, is more likely to be present bilaterally.

(Winslow, 2012)
Breast cancer is a heterogeneous disease with diverse, molecular, genetic, phenotypic, and pathologic changes. Tumor heterogeneity results from the genetic, epigenetic, and microenvironmental influences (selective pressure) that tumor cells undergo during cancer progression. Cellular subpopulations from different sections of the same tumor vary in many ways including growth rate, immunogenicity, ability to metastasize, and drug response, demonstrating significant heterogeneity. The biological attributes of a tumor as a whole are strongly influenced by its subpopulation of cells with cellular populations communicating through paracrine or contact-dependent signaling (juxtacrine) from ligands and mediated from components of the microenvironment such as blood vessels, immune cells, and fibroblasts.

Figure 1. Histological special types of breast cancer preferentially oestrogen receptor positive. (A) Tubular carcinoma, (B) cribriform carcinoma, (C) classic invasive lobular carcinoma, (D) pleomorphic invasive lobular carcinoma, (E) mucinous carcinoma, (F) neuroendocrine carcinoma, (G) micropapillary carcinoma, (H) papillary carcinoma, (I) low grade invasive ductal carcinoma with osteoclast-like giant cells. (Weigelt, Geyer, & Reis-Filho, 2010)

Figure 2. Histological special types of breast cancer preferentially oestrogen receptor negative. (A) Adenoid cystic carcinoma, (B) secretory carcinoma, (C) acinic-cell carcinoma, (D) apocrine carcinoma, (E) medullary carcinoma, (F) metaplastic carcinoma with heterologous elements, (G) metaplastic carcinoma with squamous metaplasia, (H) metaplastic spindle cell carcinoma, (I) metaplastic matrix-producing carcinoma. (Weigelt, Geyer, & Reis-Filho, 2010)
Gene expression profiling studies have identified major subtypes classified as luminal A, luminal B, HER2+, basal-like, Claudin-low, and normal breast. These subtypes have different prognoses and responses to therapy. Tumors can be stratified with gene expression profiles such as Oncotype Dx, Prosigna, and MammaPrint on the basis of genetic profiles. This information helps personalize breast cancer treatment and determine which women need aggressive systemic treatment for high-risk cancers versus close surveillance for indolent tumors.
Many models of breast carcinogenesis have been suggested and the expanding themes include (1) gene addiction, (2) phenotype plasticity, (3) cancer stem cells, (4) hormonal outcomes affecting cell turnover of mammary epithelium, stem cells, extracellular matrix, and immune function.
Cancer gene addiction includes oncogene addiction, whereby these driver genes play key roles in breast cancer development and progression. In non-oncogene addiction, these genes may not initiate cancer but play roles in cancer development and progression. Examples of key driver genes include HER2 and MYC, and examples of tumor-suppressor genes include TP53, BRCA1, and BRCA2. Once a founding tumor clone is established, genomic instability may assist through the establishment of other subclones and contribute to both tumor progression and therapy resistance.
Phenotypic plasticity is exemplified by a distinctive phenotype called epithelial-to-mesenchymal transition (EMT). EMT is involved in the generation of tissues and organs during embryogenesis, is essential for driving tissue plasticity during development, and is hijacked during cancer progression. The EMT-associated programming is involved in many cancer cell characteristics, including suppression of cell death or apoptosis and senescence. It is reactivated during wound healing and is resistant to chemotherapy and radiation therapy. Remodeling or reprogramming of the breast during post-pregnancy involution is important because it involves inflammatory and “wound healing-like” tissue reactions known as reactive stroma or inflammatory stroma. The reactive stroma releases various signals and interleukins that affect nearby carcinoma cells, inducing these cells to activate their previously silent EMT programs. The activation is typically reversible (i.e., plasticity), and those EMT programs may revert through mesenchymal-epithelial (MET) to the previous phenotypic state before the induction of the EMT program. Reactive stroma increases the risk for tumor invasion and may facilitate the transition of carcinoma in situ to invasive carcinoma. Activation of an EMT program during cancer development often requires signaling between cancer cells and neighboring stromal cells. In advanced primary carcinomas, cancer cells recruit a variety of cell types into the surrounding stroma. Overall, increasing evidence suggests that interactions of cancer cells with adjacent tumor-associated stromal cells induce malignant phenotypes.
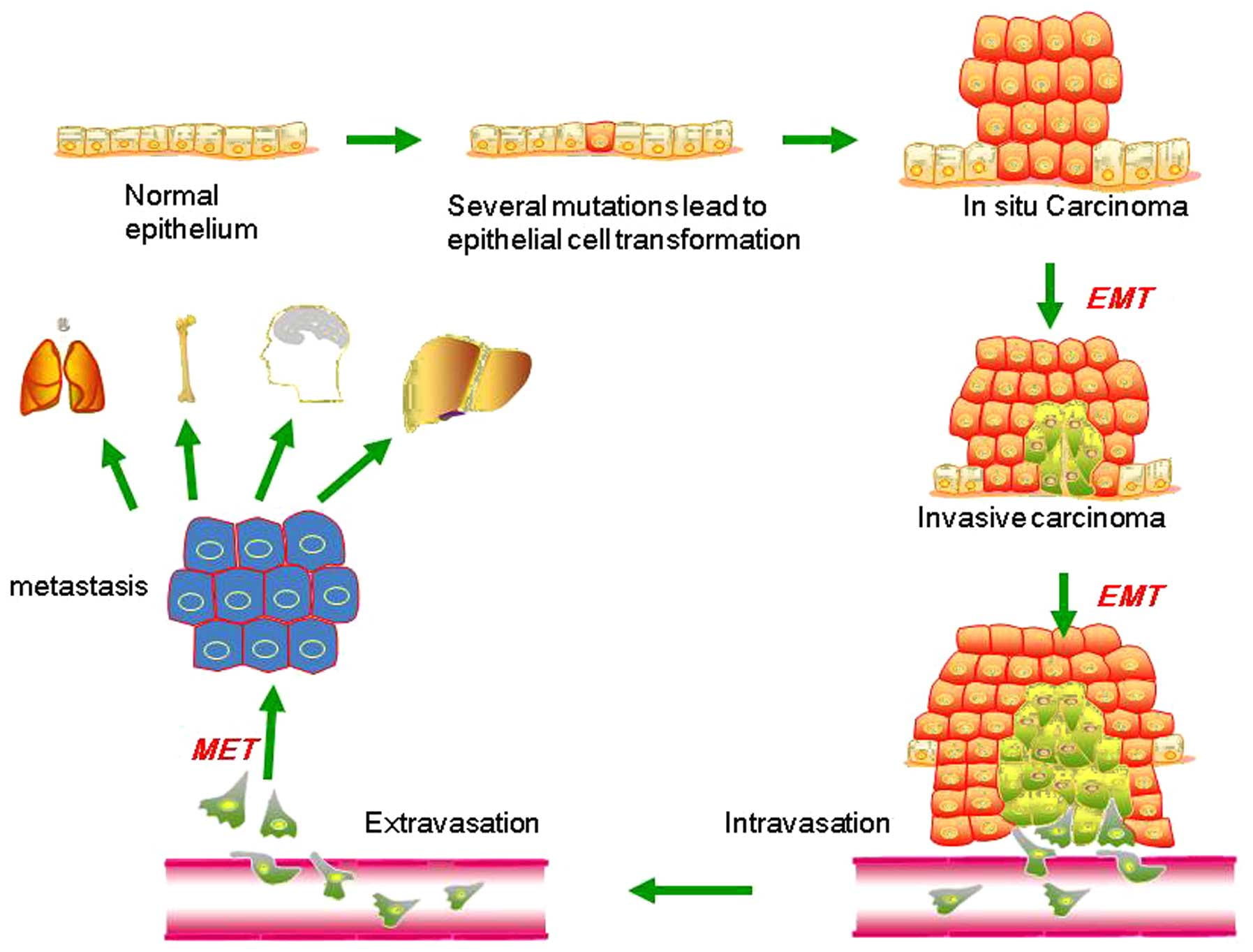
Figure 3. Putative EMT and MET in breast cancer progression. Normal epithelial cells undergo a series of transformational changes to become malignant tumor cells. Clonal proliferation of malignant cells gives rise to invasive carcinoma. Some of these cells undergo EMT and enter into the neighboring blood vessels or lymphatic vessels. These cells may remain in the circulation as circulating tumor cells or may extravasate at a distant site. The extravasated tumor cells form macrometastasis by a reverse mechanism known as MET. EMT, epithelial-mesenchymal transition; MET, mesenchymal-epithelial transition. (Liu, Gu, Shan, Geng, & Sang, 2016)
Using a mouse model of tumor heterogeneity, investigators demonstrated different clones within the heterogeneous population had distinct properties, such as the ability to dominate the primary tumor, or to contribute to metastatic populations, or to enter the lymphatic or vascular systems via vascular mimicry.

(Wagenblast et al., 2015)
Figure 4. Two adjacent sections of a mouse breast tumor. Tissue at left is stained so that normal blood vessels can be seen (brown arrow). Extending from these vessels are blood filled channels (green arrows). On the right, the tissue is stained for a fluorescent protein expressed by the tumor cells. Here it is seen that blood-filled channels are actually formed by tumor cells in a process known as vascular mimicry. The team demonstrate that the tumor cells lining these channels help drive metastasis, the process by which tumors spread. (Ravindran, 2019)
Invasion by primary tumor cells typically involves the collective migration of large cohesive groups into adjacent tissue rather than the scattering of individual carcinoma cells. However, still unknown are the precise events occurring at the invasive stage. Dormant carcinoma cells called minimal residual disease (MRD) appear to perpetuate carcinogenesis and form the precursors of eventual metastatic relapse and, sometimes, rapid cancer recurrence. Dormant cells have exited the cell cycle and are not proliferating. Thus current treatments that preferentially kill proliferating cells render dormant cells intrinsically more resistant and may remain after initial chemotherapy, radiotherapy, and surgery.
Emerging evidence supports three main prerequisites that must be met for metastatic colonization to succeed: the capacity to seed and maintain a population of tumor-initiating stem cells; the ability to create adaptive, organ-specific colonization programs; and the development of a supportive microenvironmental niche. Metastases may also occur early in the process of neoplastic transformation.

Figure 5. Extracellular matrix (ECM) changes in breast cancer progression and metastasis. The primary components of the ECM in normal mammary gland are significantly changed in breast cancer. A desmoplastic reaction is associated with breast cancer development, due to the increased production of fibrous ECM by activated fibroblasts and cancer cells. The increased collagen deposition and crosslinking by lysyl oxidase (LOX) enzymes, together with the increased production of fibronectin and other ECM components, stiffens the ECM, which in turn promotes tumor aggressiveness. The basement membrane surrounding the mammary gland epithelium is broken down by ECM remodeling enzymes like MMPs, heparanase and others. Matricellular proteins that promote cancer cell fitness such as tenascin C, periostin, osteopontin, SPARC and thrombospondin-1 are also upregulated. Breast cancer cells from the primary tumor, that include cells with the ability to establish metastatic colonies, enter the blood circulation, disseminate and can reach distant sites. While the vast majority of disseminated cancer cells are eliminated or undergo dormancy due to the adverse environment, few cancer cells are able to resist the selective pressure and establish a metastatic colony. These cells may rely on signals from the ECM such as type I collagen (collagen I), crosslinked by LOX. Tenascin C (TNC) and periostin (POSTN), which are crucial ECM proteins of the metastatic niche, promote stem/progenitor pathways and metastatic fitness in disseminated breast cancer cells. (Insua-Rodríguez & Oskarsson, 2016)
The first clinical manifestation of breast cancer is usually a small, painless lump in the breast. Other manifestations include palpable lymph nodes in the axillae, dimpling of the skin, nipple and skin retraction, nipple discharge, ulcerations, reddened skin, and bone pain associated with bony metastases.
Treatment is based on the extent or stage of the cancer and includes surgery, radiation, chemotherapy, hormone therapy, and biologic therapy.
(Unless otherwise cited, all pathophysiology information was gathered from Danhausen, Phillippi, & McCance, 2019)